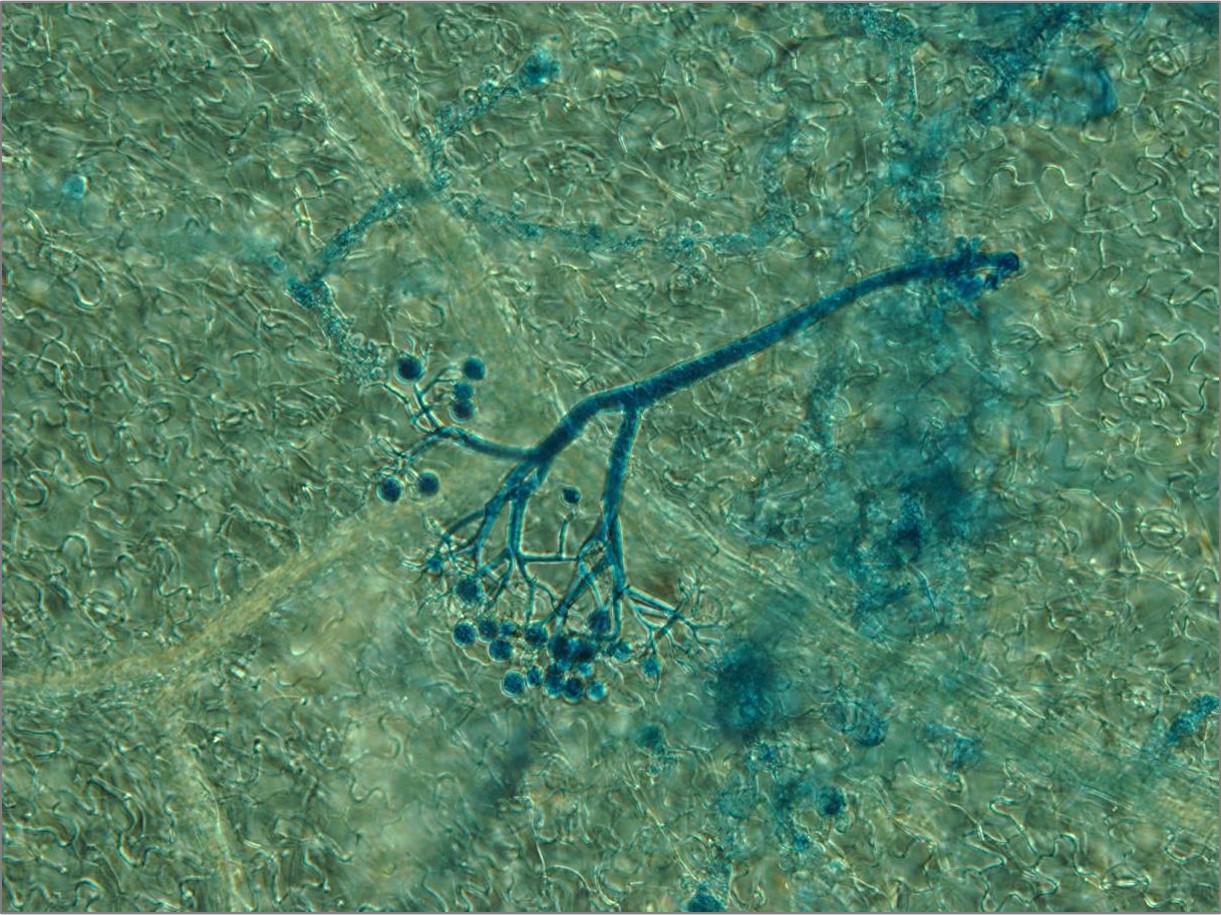
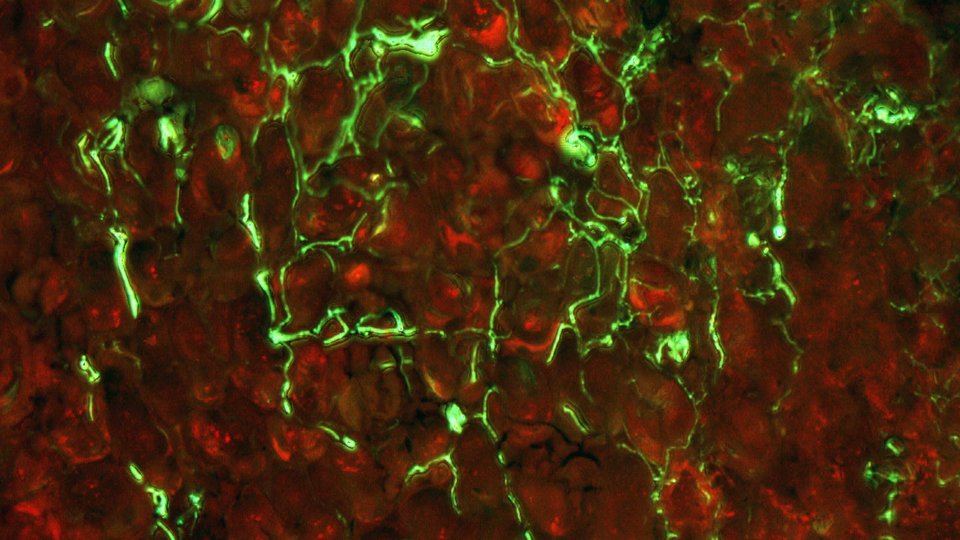
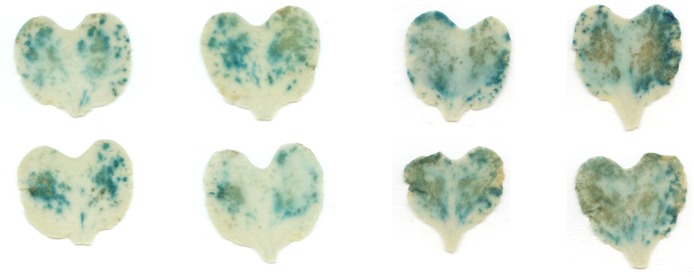
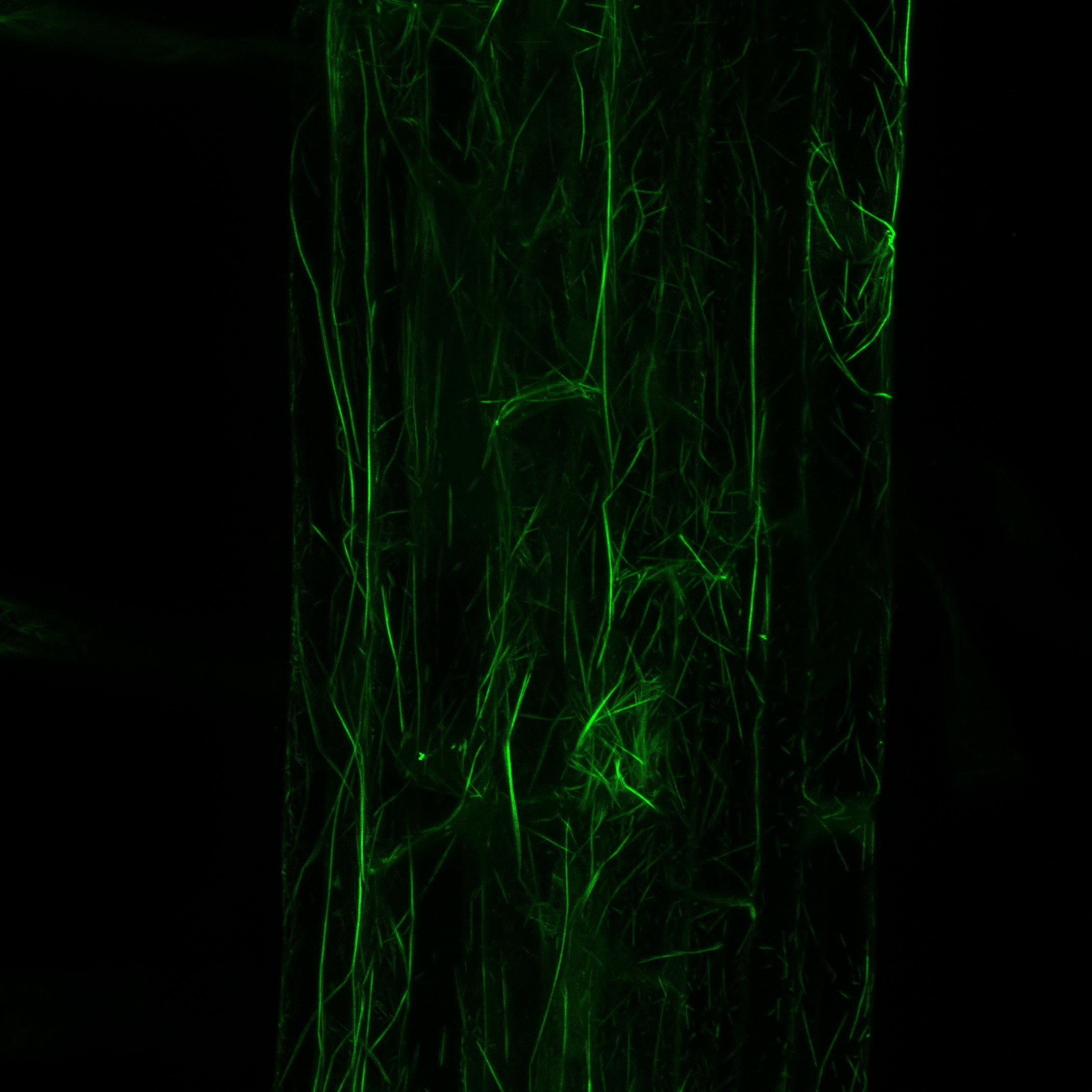
Plant-pathogen interactions are largely dependent on communication between them. Upon interaction, pathogens secrete small molecules (mostly proteins) at the infection site called effectors or factors of virulence. Typical function of effectors is to suppress plant defences.
The mechanisms of how effectors suppress plants immunity and contribute to the infection development are up to date known for just a few effectors of fungal pathogens. On the other hand, plants have developed abilities to identify the effector molecules and oppose them – to activate a second wave of defence processes that can lead to complete elimination of the pathogen. This is based on the avirulence proteins, specific for the recognition of a certain effector.
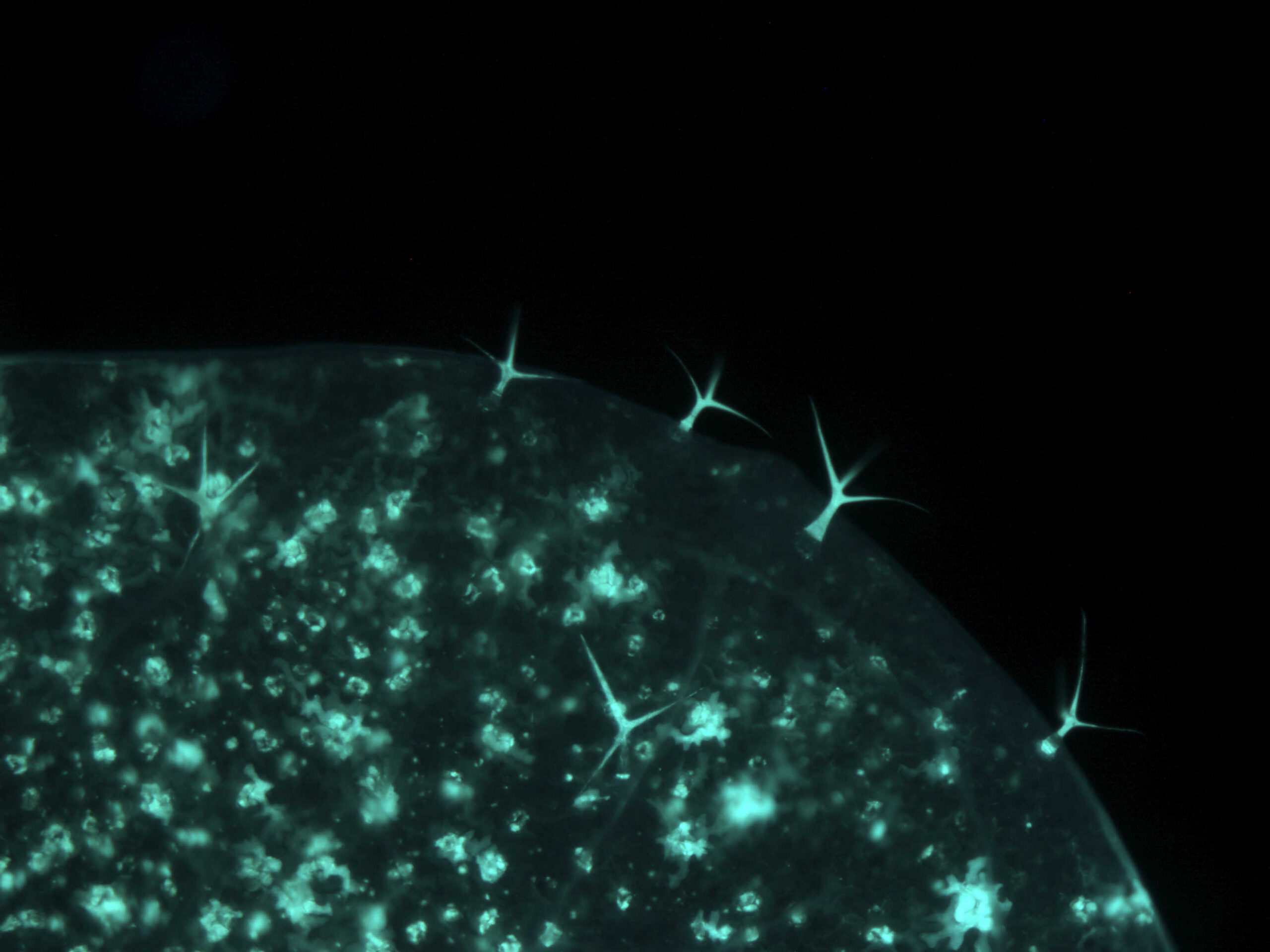
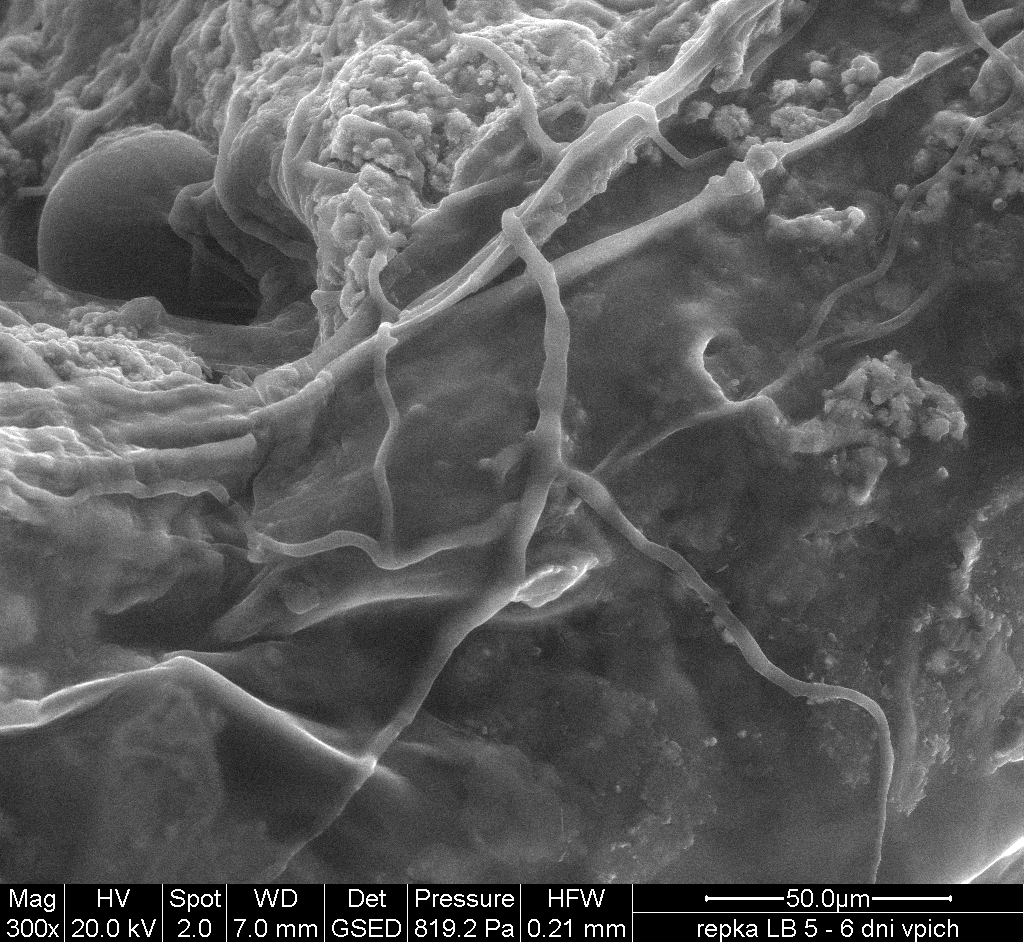
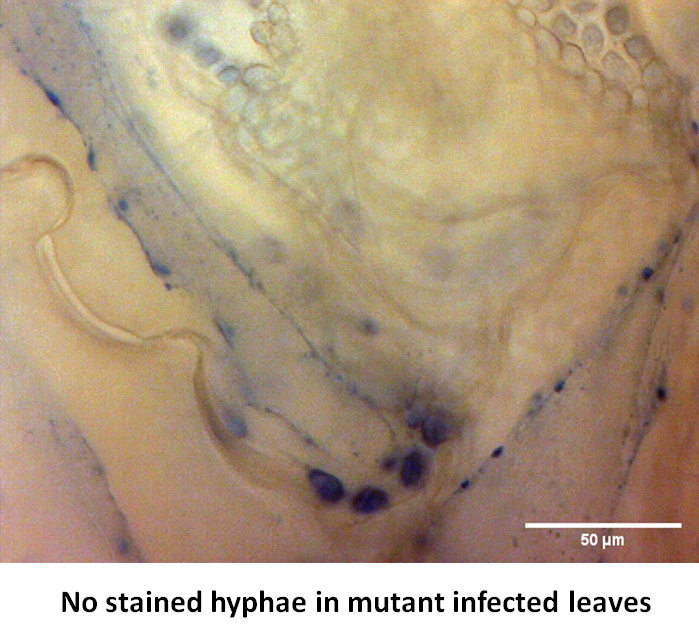
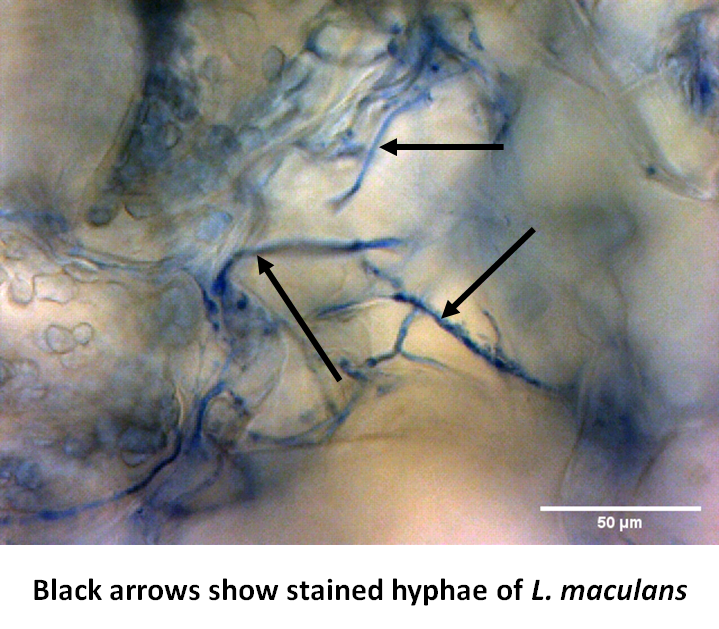
By T-DNA mutagenesis we have generated a line of L. maculans with hypovirulent phenotype which seems to be linked to a change in expression in a hexose transporter gene which we dubbed LmHXT1 and the mutant strain JN2 sp106. This phenotype is also linked to some changes in in vitro growth and mycelium morphology. On this mutant line we study mechanisms linking sugar transport and virulence.
We study the role of avirulence protein AvrLm1, which recognition activates a unique cooperation of SA, JA and ET signaling pathways with the help of reactive oxygen species (ROS). Another topic of interest is the AvrLm4-7 effector that is unique in its specificity to two plant resistance proteins and makes a remarkable contribution to L. maculans virulence. This topic also includes identification of new potential effectors or other protein molecules contributing to L. maculans virulence.